The timing of that transition, and not differential growth, is the major determinant of ultimate epidermal cell size. At the same time the cell cycle duration in the cortex gradually decreases and cells divide at a smaller size. Whether this is related to the decrease in epidermal cell proliferation and, as a result, the increased availability of a potential cell proliferation factor is an interesting question. By the end of Stage II stomata differentiation is completed. Intriguingly, the end of this stage coincides with differentiation and expansion of cortex cells, and it occurs closely after the appearance of medial vascular bundles, the final step of vasculature formation in the pedicel. These observations suggest a temporal coordination of differentiation in 4-(Benzyloxy)phenol different tissue layers during Stage II. Considering that so many changes are occurring in the epidermis and cortex during this stage, it is remarkable that the overall exponential rate of pedicel growth in Stage II is similar to the rate during Stage I. This is somewhat surprising, as the epidermis is believed to control organ growth and one might expect that multiple differentiation events in the epidermis would have an impact on the overall rate of growth. Instead, our observations are consistent with the mechanism determining the rate of organ growth being unaffected by cell differentiation in the epidermis. Stage III is a cell elongation stage and lasts for,100 hours. Very little if any cell proliferation occurs during this stage in either the epidermis or the cortex. Overall pedicel growth during this stage transitions from exponential to linear, it is not uniform along the proximodistal axis, and it depends on flower fertilization occurring just before Stage III starts. It would be most logical to compare the dynamics of pedicel growth to growth of stems, but a detailed 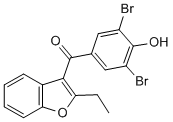 temporal analysis of stem growth is lacking. The mechanistic basis of leaf growth, however, has been studied extensively. After initiation, two stages of leaf growth are described. Epidermal and mesophyll cells initially proliferate, and then epidermal cells switch to expansion, starting at the distal end of the leaf and progressing towards the proximal end. This transition to cell expansion occurs abruptly in a specified leaf region simultaneously with the differentiation of chloroplasts and onset of photosynthesis, leading to the hypothesis that retrograde transport from chloroplasts regulates the transition to cell expansion in the epidermis. Just as in the cortex of the pedicels, the transition to cell expansion is significantly delayed in the mesophyll compared to the epidermis. Since most chloroplast differentiation and photosynthesis occurs in the mesophyll, it is somewhat surprising that a retrograde signal would induce epidermal but not mesophyll cell expansion. Stomata differentiation precedes pavement cell expansion by approximately one day, and Tulathromycin B continues until the leaf reaches its mature size. For the most part, the leaf growth pattern closely resembles the pattern of pedicel growth. In both cases growth begins with a cell proliferation stage, followed by a period in which stomata differentiate and pavement cells elongate, and ending with a period of mesophyll expansion. In both organs the differentiation of stomata slightly precedes elongation of pavement cells, and the transition to stomata differentiation occurs around day 9�C10. Despite the similarities, there are several differences between leaf and pedicel growth. The stomata differentiation period is significantly prolonged in leaves, and the onset of mesophyll expansion does not coincide with the termination of stomata differentiation. The extended stomata formation period is likely necessary for the establishment of a higher stomatal density in leaves compared to pedicels.
temporal analysis of stem growth is lacking. The mechanistic basis of leaf growth, however, has been studied extensively. After initiation, two stages of leaf growth are described. Epidermal and mesophyll cells initially proliferate, and then epidermal cells switch to expansion, starting at the distal end of the leaf and progressing towards the proximal end. This transition to cell expansion occurs abruptly in a specified leaf region simultaneously with the differentiation of chloroplasts and onset of photosynthesis, leading to the hypothesis that retrograde transport from chloroplasts regulates the transition to cell expansion in the epidermis. Just as in the cortex of the pedicels, the transition to cell expansion is significantly delayed in the mesophyll compared to the epidermis. Since most chloroplast differentiation and photosynthesis occurs in the mesophyll, it is somewhat surprising that a retrograde signal would induce epidermal but not mesophyll cell expansion. Stomata differentiation precedes pavement cell expansion by approximately one day, and Tulathromycin B continues until the leaf reaches its mature size. For the most part, the leaf growth pattern closely resembles the pattern of pedicel growth. In both cases growth begins with a cell proliferation stage, followed by a period in which stomata differentiate and pavement cells elongate, and ending with a period of mesophyll expansion. In both organs the differentiation of stomata slightly precedes elongation of pavement cells, and the transition to stomata differentiation occurs around day 9�C10. Despite the similarities, there are several differences between leaf and pedicel growth. The stomata differentiation period is significantly prolonged in leaves, and the onset of mesophyll expansion does not coincide with the termination of stomata differentiation. The extended stomata formation period is likely necessary for the establishment of a higher stomatal density in leaves compared to pedicels.
Month: June 2019
Several groups performed flow cytometric analysis to screen for the expression of markers of primary vCTB
Subsequent corruption occurs in human PDGF-driven gliomas, gliomas in general or in other tumor types, our data indicates that this process may not be limited to hPDGFb-induced murine gliomas. As such, human gliomas that appear to be comprised of two or more genetically unrelated clones, or recurrent patient gliomas that lack the genetic alterations present in the originally resected tumors subjected to therapeutic stress, may be examples of these processes. Although similar processes have not been described in vivo, anin vitro line of evidence from the colon cancer cell lines suggested that one of the mechanisms of how cancer cells bypass senescence may be related to their potential for continuous “clonal diversification”, even within the what is perceived a clonal population of cancer cells, not affected by native stromal environment. The process of corruption would thus occur if the tumor microenvironment initially drive proliferation and induce the stem-like character in both progeny of the cell-of-origin and the recruited cells. In hPDGFb-driven gliomas there is evidence for both – tumor cells ![]() secrete proliferative growth factors, inflammatory cells secrete cytokines, and endothelial and inflammatory cells secrete nitric oxide that activates the Notch signaling pathway, promoting the stem-like character. Such environment affects cells within the tumor regardless of whether they are derived from the glioma-initiating cell-of-origin or not. This notion is further supported by the analysis of expression array data using the Folinic acid calcium salt pentahydrate bacTRAP olig2 RP-eGFP reporter system and immunostaining, which suggest that recruited olig2 cells can acquire expression of GFAP and vimentin and increase the expression of nestin as compared to the tumor olig2 cells, triple co-expression of which is considered to be typical for the bona fide neural stem cells. Human placenta is a unique organ associated with fetomaternal circulation, which involves decidua basalis as the maternal component and chorionic villi from the fetus. Placenta consists of trophoblasts, which exhibit several functions such as protection, nutrition and respiration of fetus, as well as hormone production. Placental trophoblasts include relatively undifferentiated villous cytotrophoblast, intermediate cytotrophoblast, terminally differentiated villous syncytiotrophoblast and extravillous cytotrophoblast that invade into maternal decidua. These differentiated trophoblasts arise from a putative trophoblast stem cell population; it has been proposed that vCTB at the villous Lomitapide Mesylate basement membrane contains a TS cell population. In a previous study, mouse TS cell lines were established using blastocysts and extraembryonic ectoderm of E6.5 embryos cultured in vitro. They are self-renewable in the presence of FGF4 and feeder cells, and then readily differentiate into diverse trophoblast cell lineages in the absence of FGF4 and feeder cells. Human TS cell lines cannot be established under conditions similar to that used for mouse TS cell lines. The differentiation of human embryonic stem cells into trophoblasts has been studied under certain conditions. Most of the studies failed to show induction of CDX2, EOMES or ERRB in the trophoblasts, so it is controversial whether they are real human TS cells. Only a few groups presented CDX2 positive cells as putative human TS cell compartments. vCTB contains progenitor cells, and possibly, TS cells that continue to produce daughter cells that differentiate and fuse with syncytium or differentiate into invasive trophoblasts. vCTB cells can be isolated from human villous tissue at any stage of pregnancy for primary culture.
secrete proliferative growth factors, inflammatory cells secrete cytokines, and endothelial and inflammatory cells secrete nitric oxide that activates the Notch signaling pathway, promoting the stem-like character. Such environment affects cells within the tumor regardless of whether they are derived from the glioma-initiating cell-of-origin or not. This notion is further supported by the analysis of expression array data using the Folinic acid calcium salt pentahydrate bacTRAP olig2 RP-eGFP reporter system and immunostaining, which suggest that recruited olig2 cells can acquire expression of GFAP and vimentin and increase the expression of nestin as compared to the tumor olig2 cells, triple co-expression of which is considered to be typical for the bona fide neural stem cells. Human placenta is a unique organ associated with fetomaternal circulation, which involves decidua basalis as the maternal component and chorionic villi from the fetus. Placenta consists of trophoblasts, which exhibit several functions such as protection, nutrition and respiration of fetus, as well as hormone production. Placental trophoblasts include relatively undifferentiated villous cytotrophoblast, intermediate cytotrophoblast, terminally differentiated villous syncytiotrophoblast and extravillous cytotrophoblast that invade into maternal decidua. These differentiated trophoblasts arise from a putative trophoblast stem cell population; it has been proposed that vCTB at the villous Lomitapide Mesylate basement membrane contains a TS cell population. In a previous study, mouse TS cell lines were established using blastocysts and extraembryonic ectoderm of E6.5 embryos cultured in vitro. They are self-renewable in the presence of FGF4 and feeder cells, and then readily differentiate into diverse trophoblast cell lineages in the absence of FGF4 and feeder cells. Human TS cell lines cannot be established under conditions similar to that used for mouse TS cell lines. The differentiation of human embryonic stem cells into trophoblasts has been studied under certain conditions. Most of the studies failed to show induction of CDX2, EOMES or ERRB in the trophoblasts, so it is controversial whether they are real human TS cells. Only a few groups presented CDX2 positive cells as putative human TS cell compartments. vCTB contains progenitor cells, and possibly, TS cells that continue to produce daughter cells that differentiate and fuse with syncytium or differentiate into invasive trophoblasts. vCTB cells can be isolated from human villous tissue at any stage of pregnancy for primary culture.
The reduction intake was not due to lower body mass were programmed to have this alimentary behavior having had reduced energy availability
Closely a primary infection of DC in vivo than other infection models. CBAs act by binding to DENV glycoproteins and subsequently interrupt the interaction between DENV and DC-SIGN. Our data provide more insight into the mechanism of action of the CBAs in MDDC and indicate the relevance of the carbohydrate-dependent entry pathway of DENV in primary human cells. It is important to further develop therapeutic concepts that may prevent DENV-induced diseases. Small-size non-peptidic analogues, such as PRM-S, should be further pursuit for this purpose. Undernutrition is a worldwide public health issue affecting more than one billion people, particularly in underdeveloped countries, where 25% of the undernourished population is children. Therefore, several recent studies have sought to correlate undernutrition in critical periods of development with various diseases in adulthood. Studies supporting the programming hypothesis have demonstrated that adverse fetal or neonatal environments such as undernutrition result in adaptative responses leading to structural and molecular alterations in various organs and tissues. The persistence of these modifications results in the development of several diseases in adult life, particularly affecting the cardiovascular and renal systems. These pathological situations are frequently associated with hypertension. The precise mechanism involved in increased blood pressure as a late consequence of metabolic programming is as yet unclear. Most experimental data indicate that hypertension is multifactorial and involves alterations in various organs including the kidney. Kidneys play a major role in the long-term control of arterial blood pressure by regulating Na intake/excretion. It has been reported that offspring from rats that are protein-restricted throughout gestation present with marked oligonephroenia, which can lead to a reduction in pressure natriuresis and consequent elevation of blood pressure. However, the reduced number of nephrons is not the sole cause of hypertension in the protein restriction model of programming. The intrarenal molecular machinery may also be altered, contributing to the programming of hypertension. This narrow period of pup development is considered one of the most important for the development of metabolic programming, and nephrogenesis is completed during this period. Fig. 1 demonstrates that despite the increased demand of lactation from an equal number of pups, the alimentary rhythm of  undernourished mothers entailed two restrictions, low protein content of the diet and lower total food Pimozide intake with decreased available energy, which could cause metabolic programming in the offspring. The abnormal alimentary habit of undernourished mothers during lactation is probably due to low protein-associated hyperleptinemia, which could lead to satiety despite an increased energy demand as lactation ensued. The accompanying Mechlorethamine hydrochloride hypoprolactinemia contributed to the possible anorexigenic hormonal balance due to the orexigenic actions of prolactin. The reduced body mass of the offspring at weaning was a result of the low-protein induced lactogenesis failure. These maternal alimentary restrictions could contribute to the imprinting of metabolic programming of the progeny as they consistently consumed less of the normal diet despite it being offered ad libitum. This decreased food intake led to a reduced body mass throughout the growth period analyzed in the present study. Since the offspring food intake/body mass ratio after weaning remained permanently lower.
undernourished mothers entailed two restrictions, low protein content of the diet and lower total food Pimozide intake with decreased available energy, which could cause metabolic programming in the offspring. The abnormal alimentary habit of undernourished mothers during lactation is probably due to low protein-associated hyperleptinemia, which could lead to satiety despite an increased energy demand as lactation ensued. The accompanying Mechlorethamine hydrochloride hypoprolactinemia contributed to the possible anorexigenic hormonal balance due to the orexigenic actions of prolactin. The reduced body mass of the offspring at weaning was a result of the low-protein induced lactogenesis failure. These maternal alimentary restrictions could contribute to the imprinting of metabolic programming of the progeny as they consistently consumed less of the normal diet despite it being offered ad libitum. This decreased food intake led to a reduced body mass throughout the growth period analyzed in the present study. Since the offspring food intake/body mass ratio after weaning remained permanently lower.
Laboratory-adapted strains and low-passage clinical isolates, evaluated in primary MDDC
We observed neither in Raji/DC-SIGN cells nor in MDDC antiviral Cinoxacin activity of AH against DENV. This indicates that AH interacts rather specifically with high-mannose N-glycans on HIV-1 glycoprotein gp120, but not with DENV glycoprotein E. The mAb 2G12 specifically recognizes a cluster of highmannose-type oligosaccharides on HIV-1 gp120. mAb 2G12 could inhibit HIV binding to Raji/DC-SIGN + cells and could also bind to yeast glycoproteins. We therefore assumed that mAb 2G12 could potentially recognize the E-protein of DENV, however no inhibitory effect on DENV was observed. We can thus conclude that not all CBAs interact with all types of glycosylated enveloped viruses. The lectins HHA, GNA and UDA have a broad spectrum antiviral activity against HIV, SIV, HCV, HCMV and DENV but not against parainfluenza-3, vesicular stomatitis virus, respiratory syncytial virus or herpes simplex virus. This may be because of differences in carbohydrate structures on the glycoproteins of the viral envelope of different viruses grown in different host cells. The glycosylation pattern in DENV differs from HIV because they replicate in mosquito cells and human cells, respectively. In vertebrate and invertebrate hosts the glycosylation process is fundamentally different. N-glycosylation in mammalian cells is often of the complex-type because a lot of different processing enzymes could add a diversity of monosaccharides. Glycans produced in insect cells are far less complex, because of less diversity in processing enzymes and usually contain more high-mannose and pauci-mannose-type glycans. When DENV is captured by DC, a maturation and activation process occurs. DC require downregulation of C-type lectin receptors, upregulation of costimulatory molecules, chemokine receptors and enhancement of their APC function 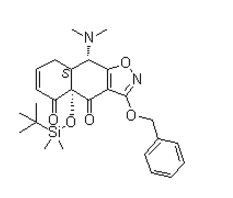 to migrate to the nodal T-cell areas and activate the immune system. Cytokines implicated in Chloroquine Phosphate vascular leakage are produced, the complement system becomes activated and virus-induced antibodies can cause DHF via binding to Fc-receptors. Several research groups demonstrated maturation of DC induced by DENV infection. Some groups made segregation in the DC population after DENV infection, the infected DC and the uninfected bystander cells. They found that bystander cells, in contrast to infected DC, upregulate the cell surface expression of costimulatory molecules, HLA and maturation molecules. This activation is induced by TNF-a and IFN-a secreted by DENVinfected DC. We observed an upregulation of the costimulatory molecules CD80 and CD86 and a downregulation of DC-SIGN and MR on the total DC population following DENV infection. This could indicate that the DC are activated and can interact with naive T-cells and subsequently activate the immune system resulting in increased vascular permeability and fever. When we examined the effect of the CBAs on the expression level of the cell surface markers of the total DC population, we are able to inhibit the activation of all DC caused by DENV and keeping the DC in an immature state. Furthermore, DC do not express costimulatory molecules and can not interact nor activate T-cells. An approach to inhibit DENVinduced activation of DC may prevent the immunopathological component of DENV disease. Immunoglobulin G was previously shown to inhibit the differentiation and maturation of DC in vitro indicating that the DC activation process is an important target for controlling immune responses in several diseases. In conclusion, we observed broad spectrum antiviral activity of HHA, GNA and UDA against all four serotypes of DENV.
to migrate to the nodal T-cell areas and activate the immune system. Cytokines implicated in Chloroquine Phosphate vascular leakage are produced, the complement system becomes activated and virus-induced antibodies can cause DHF via binding to Fc-receptors. Several research groups demonstrated maturation of DC induced by DENV infection. Some groups made segregation in the DC population after DENV infection, the infected DC and the uninfected bystander cells. They found that bystander cells, in contrast to infected DC, upregulate the cell surface expression of costimulatory molecules, HLA and maturation molecules. This activation is induced by TNF-a and IFN-a secreted by DENVinfected DC. We observed an upregulation of the costimulatory molecules CD80 and CD86 and a downregulation of DC-SIGN and MR on the total DC population following DENV infection. This could indicate that the DC are activated and can interact with naive T-cells and subsequently activate the immune system resulting in increased vascular permeability and fever. When we examined the effect of the CBAs on the expression level of the cell surface markers of the total DC population, we are able to inhibit the activation of all DC caused by DENV and keeping the DC in an immature state. Furthermore, DC do not express costimulatory molecules and can not interact nor activate T-cells. An approach to inhibit DENVinduced activation of DC may prevent the immunopathological component of DENV disease. Immunoglobulin G was previously shown to inhibit the differentiation and maturation of DC in vitro indicating that the DC activation process is an important target for controlling immune responses in several diseases. In conclusion, we observed broad spectrum antiviral activity of HHA, GNA and UDA against all four serotypes of DENV.
This is consistent with the ubiquitous dependently on the core cell death proteins
It is not clear whether additional component may exist to promote the cell killing process upstream of CED-3. Moreover, some cell death effectors that act downstream of CED-3, such as CED-8 and WAH-1, or are CED-3 substrates, such as DCR-1, are important for the timing or progression of programmed cell death. The eukaryotic translation initiation factor 3 plays Folinic acid calcium salt pentahydrate essential roles in the initiation of translation. The mammalian eIF3 complex contains 10�C13 subunits, including five highly conserved core subunits and five to eight less conserved non-core subunits. The 28 kDa human eIF3k protein was originally identified as a non-core subunit of the eIF3 complex. An in vitro reconstitution experiment showed that eIF3k is not required for the formation of the active eIF3 complex. Interestingly, eIF3k is conserved among metazoans, including C. elegans, D. melanogaster, M. musculus, and H. sapiens, but is absent in S. cerevisiae, suggesting a specialized role for eif-3.K in multicellular organisms. In addition, human eIF3k is associated with dynein, cyclin D3, the 5-HT7 receptor, and keratin K18, suggesting the involvement of eIF3k in processes that are unrelated to translation. Recently, we reported an apoptosis-promoting function for eIF3k in simple epithelial cells. Upon apoptotic stimuli, keratin K18 is cleaved by caspase 3, resulting in the collapse of K8/K18 intermediate filaments into apoptotic bodies and the sequestration of caspase 3 in kerain-containing inclusions. eIF3k binds to keratin inclusions, which in turn leads to the release of keratin-associated caspase into the cytosol to facilitate the execution of apoptosis. Keratin K8/K18 is the major intermediate filament in epithelial cells. 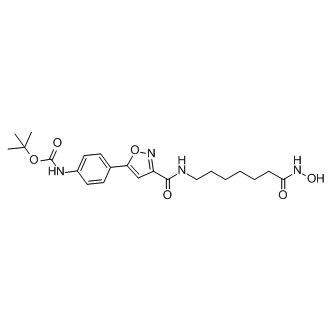 It is not clear whether eIF3k may potentiate Mechlorethamine hydrochloride apoptosis in other cell types, such as neurons or muscle cells, where intermediate filaments other than keratin are present. In addition, it is unclear whether the apoptosispromoting function of eIF3k has been conserved throughout evolution. In this work, we characterized the function of eif-3.K in C. elegans and showed that its apoptosis-promoting function has indeed been conserved throughout evolution. Furthermore, we identified a new function for the WH domain of EIF-3.K in the promotion of programmed cell death. We have previously shown that human eIF3k promotes apoptosis in cultured simple epithelial cells. In this work, we provide evidence that eif-3.K has a cell deathpromoting function at an organismal level and that this function has been conserved through evolution. In C. elegans, the loss of eif-3.K caused reduced programmed cell death and enhanced cell survival in sensitized mutants. In contrast, the overexpression of eif-3.K by the heat shock promoter or a touch neuron-specific promoter resulted in ectopic cell death. These results demonstrate that eif-3.K promotes programmed cell death. Our results also show that eif-3.K is essential for the efficient cell death that is induced by the overexpression of egl-1 or ced-4, but not ced-3, as the loss of eif-3.K partially suppresses the cell death that is induced by the overexpression of egl-1 or ced-4 only. In addition, the observation that ced-3 overexpression can rescue the cell death-defective phenotype of eif-3.K mutants and that the ced-3 strong mutation can suppress cell death caused by heat shock-induced eif-3.K overexpression further reinforces the notion that eif-3.K requires ced-3 to promote programmed cell death. Furthermore, the wide range in the identity and type of extraneous surviving cells that are affected by the eif-3.K mutation suggests that eif-3.K may be involved in the majority of programmed cell death.
It is not clear whether eIF3k may potentiate Mechlorethamine hydrochloride apoptosis in other cell types, such as neurons or muscle cells, where intermediate filaments other than keratin are present. In addition, it is unclear whether the apoptosispromoting function of eIF3k has been conserved throughout evolution. In this work, we characterized the function of eif-3.K in C. elegans and showed that its apoptosis-promoting function has indeed been conserved throughout evolution. Furthermore, we identified a new function for the WH domain of EIF-3.K in the promotion of programmed cell death. We have previously shown that human eIF3k promotes apoptosis in cultured simple epithelial cells. In this work, we provide evidence that eif-3.K has a cell deathpromoting function at an organismal level and that this function has been conserved through evolution. In C. elegans, the loss of eif-3.K caused reduced programmed cell death and enhanced cell survival in sensitized mutants. In contrast, the overexpression of eif-3.K by the heat shock promoter or a touch neuron-specific promoter resulted in ectopic cell death. These results demonstrate that eif-3.K promotes programmed cell death. Our results also show that eif-3.K is essential for the efficient cell death that is induced by the overexpression of egl-1 or ced-4, but not ced-3, as the loss of eif-3.K partially suppresses the cell death that is induced by the overexpression of egl-1 or ced-4 only. In addition, the observation that ced-3 overexpression can rescue the cell death-defective phenotype of eif-3.K mutants and that the ced-3 strong mutation can suppress cell death caused by heat shock-induced eif-3.K overexpression further reinforces the notion that eif-3.K requires ced-3 to promote programmed cell death. Furthermore, the wide range in the identity and type of extraneous surviving cells that are affected by the eif-3.K mutation suggests that eif-3.K may be involved in the majority of programmed cell death.