Fifty cells were examined for colocalization of vimentin and SERT following 5HT stimulation. In all YFP-SERT transfected cells, a limited but consistent co-localization between vimentin and SERT on the plasma Homatropine Bromide membrane was seen. The colocalization of endogenously expressed vimentin and transiently expressed SERT was monitored in 5HT-stimulated CHO- cells using IF microscopy. The cellular distribution of vimentin was significantly different in 5HTstimulated cells than in control cells. To facilitate a comparison between the localization of SERT and vimentin, the SERT signal was pseudocolored in green in merged images. 5HT stimulation mostly located vimentin around the plasma membrane. Exposure of CHO-YFP-hSERT to 5-HT induced the spatial reorientation of vimentin filaments. In control cells, vimentin exhibited a curved filamentous appearance. Vimentin filaments became more straight and bundled 30 min after stimulation with 100 mM 5HT. To evaluate the association of vimentin and the Atropine sulfate truncated and mutant transporters on the plasma membrane, the second half of the same biotinylated samples were subjected to immunoblot analysis with vimentin-Ab. In untransfected cells our W/ B analysis recognized the endogenous vimentin as one of the proteins pulled by the biotinylated plasma membrane-bound proteins. Therefore, it is clear that vimentin had bound other plasma membrane proteins as well as SERT. In SERT transfected cells, the level of vimentin on the plasma membrane was much higher than the untransfected ones. This finding identifies SERT as one of the membrane-bound proteins that links vimentin to the plasma membrane. Similarly, the levels of vimentin on the plasma membrane of D26, D20, D14, and DDD transfected cells were the same as that on the plasma membrane of untransfected cells. This finding suggests a lack of association between vimentin and D26, D20, D14, or DDD on the plasma membrane. On the other hand, the vimentinbinding abilities of T616D and T613D on the plasma membrane were very similar to CHO-SERT and CHO-D6 cells. The levels of vimentin on the plasma membrane of S611D transfected cells was lower than that in wild-type transfected cells but higher than that in untransfected ones. These findings are in good 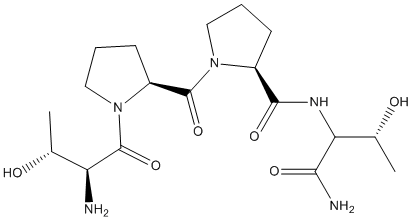 agreement with the data in Figure 6. Collectively, they support our hypothesis that the density of transporter on the plasma membrane.
agreement with the data in Figure 6. Collectively, they support our hypothesis that the density of transporter on the plasma membrane.
Month: June 2019
What spanning the cell wall and transiently expressed on cell surface
In this scenario, the antibodyaccessible b-1,3 glucan could be mostly represented by the glucan bound to the proteins of the outer cell wall, which is likely to undergo even marked variations during growth and morphologic changes, rather than the more invariant b-glucan of the inner cell wall skeleton. In this paper, we show that there is at least as much IgM- as IgG mAb-reactive b-glucan in C. albicans cell wall, as seen by ELISA using C. albicans b-glucan and EM of goldimmunolabelled sections of both yeast and hyphal cells of the fungus. Remarkably, however, the material secreted by growing fungal cells was much more reactive with the IgG than with the IgM mAb or substantially non reactive with the IgM antibody. This suggests that the secreted material is enriched with b1,3 glucan sequences as compared with b-glucan “stably” located in the cell wall. Among the secretory components recognized by the protective IgG mAb, two of them, Als3 and Hyr1, could be among the targets of the protective action of this mAb. Both these proteins belong to a major category of cell wall components, the GPIanchored proteins, which are covalently linked to b-glucan and include structural constituents, enzymes, adhesins and other components playing a crucial role in cell wall organization, stress response and virulence of the fungus. Both Als3 and Hyr1 are known to be actively secreted by C. albicans : their strong binding by the IgG mAb would also suggest that they are secreted with attached b-glucan, in particular b1,3 glucan, moieties. Relatively little is known about the functional properties of Hyr1 protein. It is activated  during hyphal transition and upon exposure to macrophages or neutrophils and its expression has been shown to be under control of well-established hyphal regulatory pathways of this fungus. Overall, this protein is assumed to play a role in yeast to mycelial conversion of C. albicans. Relevant to the data shown in this study is a very recent report that members of the IFF protein family, to which Hyr1 belongs, play a role in Orbifloxacin fungus adherence. There is much more information on the structure and biological properties of Als3, and this is clearly relevant to the interpretation of the inhibitory and protective potential of our anti-b1,Labetalol hydrochloride 3-glucan IgG mAb. Als3 is encoded by the corresponding gene of the ALS gene family which code for structurally-related, high Mr cell surface and secreted glycoproteins.
during hyphal transition and upon exposure to macrophages or neutrophils and its expression has been shown to be under control of well-established hyphal regulatory pathways of this fungus. Overall, this protein is assumed to play a role in yeast to mycelial conversion of C. albicans. Relevant to the data shown in this study is a very recent report that members of the IFF protein family, to which Hyr1 belongs, play a role in Orbifloxacin fungus adherence. There is much more information on the structure and biological properties of Als3, and this is clearly relevant to the interpretation of the inhibitory and protective potential of our anti-b1,Labetalol hydrochloride 3-glucan IgG mAb. Als3 is encoded by the corresponding gene of the ALS gene family which code for structurally-related, high Mr cell surface and secreted glycoproteins.
Which are both intrinsic cell wall constituents and secretory molecules some
Microarray analyses showed exclusive IgG mAb binding to b1,3-linked glucose sequences with DP 4 and greater. In contrast, the IgM mAb bound both to b1,3- as well as to b1,6-linked glucose sequences with DP 2 and greater. As shown by microarray analyses it also recognized saccharides with b1,4 glucan sequences, which are ordinarily absent in fungi but present in higher plants and lichens. Interestingly, a linear b1,3-b1,4-linked glucan is uniquely present in the cell wall of Aspergillus fumigatus, a fungus which is bound by both mAbs, as found by immunofluorescence staining, and whose growth is highly susceptible to the anti-b-glucan antibodies generated by the immunization with the Lam-CRM vaccine. The disposition of b1,3 glucan on C. albicans cell surface, an essential Mepiroxol prerequisite for interaction with the protective anti-bglucan mAb, is a matter of controversy. There are suggestions that b1,3 glucan is hidden inside the cell wall by a layer of mannoprotein constituents and that it is lacking or negligibly Danshensu expressed on the fungal surface, particularly in hyphal cells. This sort of b-glucan stealth has been recently interpreted as a host-deceiving mechanism, as it would prevent the binding of fungal cells by Dectin-1 – a b1,3 glucan specific receptor present on a variety of cells of the innate immune system – thus inhibiting the triggering of critical signals for coupling innate to adaptive antifungal immunity. However, other authors have reported surface expression of this polysaccharide in both yeasts and hyphae. This apparent discrepancy could be reconciled by very recent reports that b1,3 glucan may be variably expressed by fungal cells depending on their growth conditions and/or morphology. Moreover, different reagents, with likely different fine specificity, were used by the various Authors to detect b1,3 glucan at cell surface. The IF and EM data shown here with the use of a mAb with restricted specificity for b1,3 glucan leave little doubt that some b1,3 glucan or components carrying this polysaccharide are expressed on cell surface, albeit not in all cells of the culture and not uniformly in all labelled cells. This would be expected in a asynchronous culture if b1,3 glucan is consistently exposed only at certain stages of growth. Interpretation of all these data must take also into account the presence of b1,3 and b1,6 glucan moieties in the mannoprotein modules of .gif) the cell wall.
the cell wall.